
Spoluator: Lukáš Hleba. Katedra mikrobiológie, Fakulty biotechnológie a potravinárstva, Slovenská poľnohospodárska univerzita v Nitre
Morfológia
Ako by sme si mohli predstaviť hubu predátora? Je to organizmus, zložený z jednotlivých ale i vzájomne prepojených mikroskopických vlákien (modulov), na ktorých sa vyskytujú pasce v podobe lepkavých výbežkov, sťahujúcich, či fixných očiek, výbežkov produkujúcich toxíny či trojrozmerné lepkavé siete tzv. modifikované moduly, ktorými huba chytá okolo sa pohybujúcu korisť. To, že sa hovorí o mikroskopických hubách je trochu zavádzajúce, pretože celková veľkosť tela huby (hýfového systému) môže byť len niekoľko málo desiatok mikrometrov ale tiež aj veľa metrov štvorcových. Na špecializovaných hýfach – nosičoch (sporangioforoch a konídioforoch) týchto húb môžeme často nájsť nepohlavné spóry, ktoré môžu byť jedno alebo viacbunkové, ktoré sú obalené mukóznou (hlienovitou) vrstvou a taktiež môžu fungovať ako pasce. Ako príklad môže poslúžiť originálne vyobrazenie Dactylella gampsospora alebo Triposporina quadridens na ktorých je vidieť hýfy, konídiofory s konídiami, prípadne i pascu s chytenou korisťou.

Obr.č.1 Dactylella gampsospora
U niektorých dravých húb je známe aj pohlavné rozmnožovanie, kedy sú tvorené misčičkovité plodničky – apothécia (napr. pri rode Orbilia), prípadne tiež bazídiokarpy (napr. pri rode Plerotus – Hliva)
Predácia mikroskopických húb
Predácia má u modulárnych organizmoch akými sú huby alebo rastliny trochu iný rozmer na aký sme zvyknutí. Ako predátor je všeobecne myslený taký organizmus, ktorý konzumuje organizmus iný alebo aspoň jeho časť. Predátory môžeme rozdeliť na pravé predátory, parazity, parazitoidy a spásače. Praví predátori korisť zabíjajú viac menej hneď po útoku a počas života zabíjajú ešte mnoho jedincov. Paraziti a parazitoidi spotrebúvajú často len časť svojej koristi a väčšinou sa počas života sústreďujú len na jedného jedinca, ktorého zabíjajú až po dlhšej dobe alebo vôbec nikdy. Spásači svoju korisť neusmrcujú a spotrebúvajú len jej časť ako napr. krava na paši alebo pijavica.
Je evidentné, že toto rozdelenie nieje celkom dokonalé a môžeme nájsť mnoho výnimiek a prechodov. Podľa tohto poňatia môžeme medzi predátory zaradiť takisto hubu Pythium (Oomycota), ktorá likviduje klíčne rastlinky a takisto vlka prenasledujúceho losa. Interakcie predátorov a ich koristí sú v oboch prípadoch veľmi zložité, zaujímavé a dynamikou (napr. Pythium môže zdolať klíčnu rastlinku omnoho skôr ako vlci losa, pokiaľ sa im to vôbec podarí) aj porovnateľné, no v podstate tak odlišné.
Unitárne organizmy (ako je napr. vlk) sa vyznačujú dopredu danou, určenou formou tela (všetci vlci majú štyri nohy, dve uši, atď.) Táto forma tela sa nemení od včasného embryonálneho vývojového štádia až do smrti organizmu (sú samozrejme výnimky – napr. hmyz, u ktorého sa vyskytujú rôzne vývojové štádiá, ale aj tieto štádiá majú opäť pevnú a nemennú stavbu. U modulárnych organizmov (rastliny, huby, polypy a i.) sa rozmnožovacie častice vyvíjajú v jednotný stavebný celok (modul), ktorý sa ďalej reprodukuje a tvorí organizmus. Výsledný počet modulov a ich zoskupenie nieje dopredu dané, a rovnako aj ich funkcia veľmi závisí od okolitého prostredia (napr. noha bude vždy noha, ale napr. asimilačná funkcia ríbezle sa zmení po zahrnutí hlinou na funkciu absorpčnú. U modulárnych organizmov je niekedy veľmi problematické definovať pojem jedinec (napr. u odnožujúcich rastlinách). Starnutie modulárnych organizmov sa môže prejavovať ako na úrovni jednotlivých modulov, tak aj na úrovni celého organizmu (niekedy je však starnutie naprogramované len na úrovni modulu a jedinec či polykormón je de facto nesmrteľný.
Dôsledkom týchto rozdielov je úplne odlišná biológia a životná stratégia oboch typov organizmov. Uzavretosť a pohyblivosť unitárnych organizmov im umožňuje napr. schopnosť vyhľadávať zdroje potravy, úniku pred nebezpečím či nepriaznivými podmienkami. Modulárne organizmy sú otvorené, viacej vystavené vplyvom prostredia a väčšinou neschopné pohybu. Ich výhodou (a nutnosťou k prežitiu) je plasticita a prispôsobivosť, ktorá im umožňuje vyrovnávať sa s veľkými zmenami faktorov prostredia a ktorú len veľmi ťažko nájdeme pri unitárnych organizmoch. Biológia modulárnych organizmov je relatívne viac mnohotvará a obtiažnejšie uchopiteľná, asi preto, že sami nie sme modulárne organizmy.
Pokiaľ je aspoň predátor unitárneho typu, je relatívne jednoduchšie posúdiť o aký typ predácie sa jedná. V prípade, kedy je korisť i predátor modulárneho typu je situácia často diskutabilnejšia a naše definície pre unitárne organizmy často zlyhávajú. V tomto prípade nám pojmy predácia a parazitizmus splývajú (je to evidentný dôsledok predovšetkým absencie pohybu modulárnych organizmov a tiež nejasného pojmu jedinec, z ktorého definícia predácie rovnako vychádza) a viac-menej sú tieto interakcie chápané ako parazitické, ako v prípade Pythium – klíčne rastlinky. Môžeme však bežne nájsť typ predácie, kedy predátor je modulárneho typu a korisť unitárneho typu (unitarita koristi nám trochu pomôže pri určovaní povahy predácie), potom sa často jedná o typický parazitizmus (napr. huba Dermatophyton verzus človek) alebo výnimočne o pravú predáciu.
Medzi typické modulárne predátory môžeme zaradiť napr. mäsožravé rastliny či dravé huby. Problémom týchto predátorov je ich absencia pohybu a s ňou súvisiaca neschopnosť vyhľadávania a chytania koristi a rovnako je jedným z najvýznamnejších dôvodov, prečo je v týchto skupinách predácia tak málo rozšírená (najdôležitejší je samozrejme fakt, že fylogeneticky pôvodne rastliny a huby získavajú živiny a ďalšie látky iným spôsobom a nie sú k predácii naprogramované). Veľkú úlohu hrá samozrejme i schopnosť úniku potenciálnej koristi. V takejto situácií ma predátor len jedinú voľbu – loviť pomocou pascí (ako napr. pavúci) a pokiaľ je možné mať aj nejakú návnadu, ktorá by zvýšila frekvenciu výskytu koristi (ako kajmanka supia alebo hlbokomorské ryby so svetielkami).
Huby paraziti príbuzní predátorom
Predátory z ríše húb a im podobné parazity (ktorí rovnako produkujú pasce a chemické atraktanty) je možné odlíšiť v základe nasledujúcim spôsobom. U predátorov sú pasce v pravom slova zmysle –odvodené z hýf huby. Tieto pasce slúžia k chytaniu koristi a pokiaľ je korisť chytená, zostáva pripútaná k mycéliu. Jeden jedinec predátora je schopný v rôznych ontogenetických fázach vývoja uloviť desiatky (niekedy aj oveľa viac) kusov koristi. Predátorom podobné parazity majú len konídie pokryté lepkavou substanciou. U parazita nieje cieľom chytenie koristi, ale prichytenie jednej alebo viac konídií (t.j. zárodok nového jedinca) na telo koristi, ktorá ich odnáša so sebou. Ide tu teda nielen o zaistenie zdroja energie ale aj o možnosť rozšírenia. Ontogenetický vývoj jedného alebo i viacerých jedincov parazita je takto naviazaný len s jedným jedincom koristi. Podobne dochádza občas i k odlomeniu pasce predátora (je to bežné napr. u fixných očiek, ktoré majú tenkú stopku) pokiaľ sa chytená korisť príliš zmieta. Takáto pasca však nestráca svoj infekčný potenciál a po zabití koristi sa na tom istom mieste predátora vytvára ďalšia pasca. V tomto prípade sa teda do značnej miery uplatňuje i zoochória. Zoochória, je rozširovanie napríklad plodov a semien prostredníctvom živočíchov, a môže mať rôzne podoby: epizoochória (na tele), endozoochória (prechodom cez tráviacu sústavu), synzoochória (napr. rastliny, ktoré vytvárajú na svojich semenách štruktúry pre lákanie živočíchov (hlavne pre mravce)). Vzhľadom k tomu, že u húb, majúcich tento typ pascí, nachádzame i ďalší typ, kde dochádza k pevnému pripútaniu koristi k mycéliu. Môžeme špekulovať o modifikácii tejto pasce behom vývoja a postupnú náhradu funkcie zariadenia zaisťujúceho energiu funkciou zaisťujúcou šírenie alebo aspoň pozoruhodnú kombináciu oboch vlastností.
Rozdeľovanie húb na parazity a predátory je samozrejme umelé a založené len na antropomorfickom pohľade. Nemá akýkoľvek praktický význam a nieje založený na fylogenetických vzťahoch, nakoniec ani v „Dictionary of The Fungi“ nie sú tieto skupiny húb oddelené a pod pojmom “predacious fungi“ nájdeme názvy tak dravých húb (Arthrobotrys, Monacrosporium, Zoopagales), ako aj celkom nepríbuzných parazitov (Harposporium a Verticillium). Ani väčšina autorov zaoberajúcich sa problematikou tejto skupiny húb striktne neodlišuje predátory a im príbuzné parazity a to hlavne z dôvodov ich blízkej fylogenetickej príbuznosti a podobnej ekológii. Ale nakoniec keď tam rozdiel je, prečo ho nepomenovať?
Ekológia dravých húb
Dravé huby môžeme nájsť najčastejšie v pôdnom humuse alebo v hnijúcom dreve či lístí, veľa z nich i vo vodnom prostredí. Bežnou korisťou predátorov sú organizmy do veľkosti zhruba 100 mikrometrov – prvoky (hlavne rôzne améby), vírnici, háďatká, niekedy chvostoskoky alebo drobné roztoče.
Drvivá väčšina predátorov nie sú vo svojej výžive striktne viazaní na korisť – jej výskyt môže byť do istej miery náhodný a hube by sa mohlo silne nevyplatiť spoliehanie sa na jej nepretržitý dostatok (alebo absencia dlhodobého nedostatku). Preto všetky huby predátory sú do väčšej či menšej miery schopní typickej saprotrofnej výživy, ktorá je u húb vývojovo pôvodná. Veľmi často jeden jedinec využíva oba spôsoby získavania energie (prevaha toho ktorého spôsobu ja závislá na prítomnosti koristi v prostredí a rovnako na prítomnosti saprofytických kompetítorov). Rada fakultatívnych predátorov využíva korisť skôr ako zdroj dusíka než ako zdroj energie – sú to huby, ktoré žijú v prostredí chudobnom na dusík, ako je napr. drevo. Typickým príkladom je Hliva ustricová.
Na vznik pascí (t.j. na prechod od saprofytickej k predačnej fáze) pôsobí viac faktorov – endogénnych a exogénnych. Medzi endogénne môžeme zaradiť dispozície určitého kmeňa huby k ich tvorbe a štádium v ontogenéze – t.j. vek mycélia. Nedávno bola rovnako doložená periodicita tvorby pascí u huby na mycéliu priamo v pôde s Arthrobotrys oligospora (Deuteromycotina) periódou okolo 29 hodín, ktorá súvisela s maximom obsahu minerálnych látok v mycéliu predátora. Exogénna regulácia je oveľa viac preskúmaná – môžeme sem zaradiť napr. nutričný status substrátu – za nedostatok ľahko využiteľných živín saprofytickou cestou je indukcia tvorby pascí vyššia. Ďalšie pôsobiace faktory sú vlhkosť, pH, svetlo, pomer O2/CO2, prítomnosť ďalších hubových kompetítorov a množstvo tekutých látok v prostredí. Ďalej sem patrí produkcia látok potenciílnou korisťou, napr. háďatká, ich prítomnosť v prostredí indukuje tvorbu pascí. Zdá sa, že najvýznamnejšie sú krátke peptidy s veľkým podielom nepolárnych a aromatických aminokyselín. Masívny prírastok v tvorbe pascí nejde vždy dobre doložiť prítomnosťou týchto látok, aj keď môžu spôsobovať indukciu už pri nízkej koncentrácii. Vysvetlením tohto faktu bolo nedávne objavenie prenosu signálu plazmatickou membránou v celom mycéliu predátora. Odpoveď huby na prítomnosť týchto látok je pomerne veľmi rýchla – jednoduché typy pascí môžu byť vytvorené i za necelú hodinu.
Morfológia pascí
Pri podrobnejšom pohľade do sveta húb sa človek často neubráni údivu nad dokonalosťou a fascinujúcou „nápaditosťou“ týchto organizmov, s akou dokážu vynájsť množstvo rôznych fungujúcich riešení jedného problému. Rovnako tomu je aj v prípade dravých húb, kde zatiaľ poznáme jedenásť základných typov pascí a o ich úspešnom používaní nemožno pochybovať. Pasce húb sa vyznačujú veľkou rôznorodosťou. Nachádzame tu napr. pasce, ktoré sú celkom iste odlišného pôvodu, aj keď ich morfológia a mechanizmus chytania koristi sú podobné (napr. lepkavé výbežky u niektorých pododdelení: Ascomycotina, Zygomycotina, alebo Basidiomycotina).
Na druhej strane sa stretávame s morfologicky i funkčne celkom odlišnými typmi pascí, ktoré sú naopak príbuzné (napr. fixné, sťahujúce očká a siete napr. v rodoch Monacrosporium, Dactylaria a Arthrobotrys). Typy pascí sú (aspoň v tomto prípade) dobre preštudované a môžu byť charakteristickým znakom, ktorý môže byť uplatnený v taxonómii. Jedinou nevýhodou občas býva postupná degenerácia pascí behom dlhodobej kultivácie týchto húb v neprirodzených podmienkach. Takisto sa môžeme stretnúť s prítomnosťou viacerých typov pascí pri jednom druhu huby (na mycéliu jedného jedinca). U niektorých (často obligátnych) predátorov môže dochádzať k tomu, že spóra klíči priamo pascou a nie hýfou a dokonca, že tieto tzv. konidiálne pasce môžu niekedy fungovať ako trvalé štádiá. V charakteristike jednotlivých pascí je uvedený ich popis, princíp fungovania, typický zástupca, u ktorého ho môžeme nájsť a prevažujúci druh koristi. Názvy pascí sú uvádzané aj v anglickom originály, pretože pre niektoré typy pascí sa nám nepodarilo nájsť vhodné slovenské ekvivalenty a tu používaný preklad vždy celkom nezodpovedá.
Prehľad typov pascí
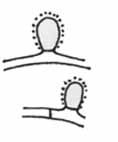
Sediaci lepkavý gombík („sessile adhezive knob“)
– je guľovitá alebo pretiahnutá bunka, pokrytá lepkavou substanciou. Niekedy tieto bunky majú tendenciu k proliferácii a potom môžu tvoriť krátke vetvy, ktoré sa niekedy zohýbajú a môžu tvoriť oblúky. Tento typ pasce je veľmi rozšírený (bežný je napr. u rodu Monacrosporium zo skupiny Deuteromycotina). Mechanizmus chytania je veľmi jednoduchý – napr. háďatko sa dotkne tohto výbežku a prilepí sa. Niekedy sa môže stať, že sa koristi podarí uniknúť, pričom často vytrhne pascu z mycélia huby a pokračuje s ním na svojom tele. Pasca však nestráca svoj infekčný potenciál a po čase dôjde k jej vyklíčeniu a následnému prerastaniu hýfy do tela koristi. V prípade, že by k tomuto javu dochádzalo bežne, by niekedy možno bolo vhodné hovoriť skôr o parazitizme a naviac by sme mohli pozorovať adaptáciu, ktorá by zjednodušovala oddelenie pasce od mycélia. Pri mnohých čisto parazitických druhoch dochádza k infekcii hostiteľa podobným spôsobom ako infekčná jednotka však slúži nepohlavná spóra (konídia), vznikajúca na diferencovanej hýfe – konídiofore. Tzv. lepkavý gombík môžeme nájsť i u niektorých druhov rodu Nematoctonus (nepohlavné štádium rodu Hohenbuehelia, Tricholomataceae, Basidiomycotina), ktorý má niektoré drobné morfologické odlišnosti a jeho „majitelia“ sú výrazne špecifickí – chytajú len niektoré druhy nematód. Funkčne podobný typ pasce je stefanocysta (biela guľovitá bunka, ktorá je na báze lemovaná vencom pozostávajúcim z ostňov).
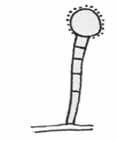
Stopkatý lepkavý gombík („stalked adhezive knob“)
– je opäť bunka, krytá lepkavou substanciou, tento krát však vynesená krátkym, niekoľko bunkovým vláknom nad mycéliom huby. Chytanie koristi prebieha rovnakým spôsobom ako u predchádzajúceho typu, len je o niečo efektívnejšie vďaka vyneseniu pasce do priestoru. Chytané môžu byť háďatká (niekedy nie príliš selektívne), ale napr. aj chvostoskoky – gombík je v tomto prípade niekoľkonásobne zväčšený a simuluje spermatofory koristi. Lepkavé stopkaté gombíky sú typické pre niektoré druhy rodov Dactylella a Monacrosporium.
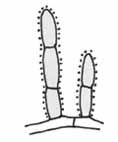
Postranné myceliálne vetvy („hyphal branches“, „adhezive columns“)
– sú krátke, niekoľko bunkové, pomerne robustné adhezívne výbežky, vyrastajúce kolmo po stranách nosnej hýfy. Často vyrastajú vedľa seba a majú tendenciu k anastomózam a môžu tvoriť jednoduché, pravouhlé vetvenia, dvojrozmerné siete. Vyskytujú sa z časti pri rode Monacrosporium.
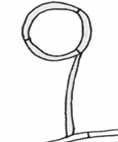
Fixné očká („non-constricting rings“)
– sú tvorené tromi bunkami na tenkom, kolmo z hýfy vyrastajúcom stĺpiku. Vyskytujú sa len u niekoľkých druhoch napr. u Dactylella leptospora a vždy spolu so stopkatými gombíkmi (nie sú totiž príliš efektívne a chýba im adhezívna vrstva). Často sú zmietajúcou sa korisťou odtrhnuté z mycélia, ale opäť si uchovávajú životaschopnosť a po čase korisť infikujú. Pomocou týchto pascí sú neselektívne chytané drobné červy.
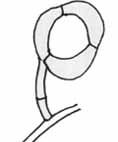
Sťahujúce očká („constricting rings“)
-sú povestné rýchlosťou pohybu – behom 0,1 sekundy bunky očka dokážu niekoľkonásobne zväčšiť svoj objem a uväzniť korisť, ktorá sa nachádza vo vnútri slučky. Je to najrýchlejší známy nedisruptívny pohyb v ríši húb. Vlastná sťahujúca slučka je tvorená kruhom troch buniek, pokrytých adhezívnou látkou, ktoré nasadajú na dvojbunkovú stopku, vyrastajúcej viac menej v pravidelných vzdialenostiach kolmo na hubových vláknach. Tieto pasce sú pomerne časté a vyskytujú sa len u obligátnych predátoroch (sú známe pri mnohých druhoch rodov Dactylella, Dactylaria, Arthrobotrys a Monacrosporium). Korisťou týchto húb sú opäť predovšetkým háďatká, ale rovnako môžu byť chytané najrôznejšie ďalšie drobné živočíchy (napr. niektorý vírnici). Niektorý predátory, používajúci sťahujúce očká, sú vysoko selektívny a zameriavajú sa len na niektoré druhy nematód. Nieje potreba dodávať, že tento typ pascí je veľmi efektívny a asi najpodivuhodnejší zo všetkých lapacích mechanizmov húb. Neobyčajnosť tohto lapacieho mechanizmu bola podnetom k mnohým štúdiám, takže dnes poznáme fungovanie tohto orgánu podrobnejšie.
(Indian Grey Matter, 2017)
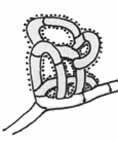
Siete („nets“)
– sú tvorené množstvom vzájomne prepojených oblúkov a slučiek. Sú najčastejšie trojrozmerné, vyčnievajú nad mycélium pokrývajúc substrát a na povrchu je lepkavá vrstva. Vyskytujú sa skôr u fakultatívnych predátorov (ktorý sú schopný rýchleho rastu aj pri čisto saprotrofnej výžive) ako doplnkový zdroj dusíkatých látok. Najčastejšou korisťou sú háďatká, ale chytať môžu široké spektrum organizmov, pretože sieťoví predátory nie sú príliš selektívni. Siete sú bežným chytacím zariadením u niektorých druhov rodu Arthrobotrys, Monacrosporium, Geniculifera a Dactylella. Mechanizmus vzniku sietí je podobný ako u fixných či sťahujúcich očiek. Základom pasce je kolmo vyrastajúca hýfa, ktorá sa bezprostredne stáča späť a zhruba po 20 – 25 mikrónoch sa približuje k drobnému výrastku na materskej hýfe, s ktorým neskôr splýva. Za presnosť tohto procesu zodpovedajú chemické atraktanty, produkované drobným výbežkom v mieste budúceho spojenia.
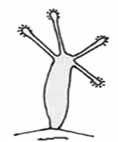
Kolíčky („pegs“)
– sú to kolíkovité výbežky, ktoré sú buď v pravidelných intervaloch na hýfach, alebo sa môžu vyskytovať v menších skupinách v terminálnej časti oválnych či kyjakovitých hrubostenných výbežkoch alebo azygospórach, vyrastajúcich kolmo na hýfach – celý útvar tak potom pripomína nezmara. Na konci týchto kolíčkov je opäť vylučovaná adhezívna látka. Najčastejšou korisťou sú vírnici, ktorý sa pokúšajú pohltiť zdurené konce týchto kolíčkov. Tento typ pascí sa vyskytuje napr. pri rode Zoophagus a Cephaliophora (u tohto rodu sú oddelené priehradkou). Sú to organizmy neistého taxonomického začlenenia – pretože skôr boli niektoré z týchto húb mylne zaradené do oddelenia Oomycota, v súčasnosti sú zaradené do pododdelenia Zygomycotina, ale niekedy je diskutovaná aj prípadná príslušnosť niektorých rodov do pododdelenia Deuteromycotina.
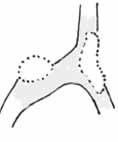
Nešpecifické výbežky („protuberances“)
– sú to akoby hýfy s vankúšmi, ktoré vylučujú adhezívnu látku, sú morfologicky prakticky nerozoznateľné od normálnych hýf predátora. Bežnou korisťou týchto húb sú jednobunkové organizmy, najčastejšie améby. Po kontakte s pascou améba zatiahne panôžky, prestane sa pohybovať a behom niekoľkých hodín je kolonizovaná hýfami dravej huby. Niekedy sú chytané tiež hlísty, vírnici a prípadne aj ďalšie organizmy. Väčšina týchto húb sú obligátny predátory – nie sú schopný získavať energiu saprotrofným spôsobom výživy. Tieto organizmy patria do oddelenia Zygomycotina a typickými zástupcami sú napr. rody Acaulopage, Zoopage, Cystopage a Stylopage. V literatúre je spomenutý tento typ pascí aj u ďalších húb so schopnosťou predácie. Z oddelenia Deuteromycotina môžeme uviesť rody Haptocara, Triposporina, Pedilospora Tridentaria, ktoré používajú k chytaniu koristi (najčastejšie améb a háďatok) buď nešpecifické adhezívne výbežky alebo polkruhové bunky s lepkavou vrstvou. Niektoré druhy rodu Tridentaria sú zaujímavé tým, že bezprostredne po chytení nematódy, začínajú z hýfy predátora blízko miesta chytenia rásť proti sebe dve vlákna, ktoré korisť pevne obopnú a spoja sa. Pre zaujímavosť – rýchlosť rastu hýf húb môže dosahovať od desiatok až do 500 mikrometrov za hodinu, čo pri nepatrnom priemeru koristi (okolo 5 – 20 mikrónov) pohodlne postačuje.

Stefanocysty
– sú jedno alebo častejšie dvojbunkové oválne útvary na mycéliu alebo na bazídiospórach niektorých druhov rodu Hyphoderma (Coticiaceae, Basidiomycotina). V ekvatoriálnej časti stefanocysty je prstenec výrastkov, ktoré sú pokryté adhezívnou vrstvou. Mechanizmus chytania je v podstate rovnaký ako u sediacich lepkavých gombíkoch. Bezprostredne potom, čo sa korisť prilepí k ostňom, stráca často stefanocysta svoj pôvodný tvar a stáva sa vakovitou, čo umožňuje lepší kontakt s korisťou a uľahčuje prenikanie infekčnej hýfy. Najčastejšou korisťou tejto huby sú háďatká, ktoré fungujú tiež skôr ako doplnkový zdroj dusíka. Ešte donedávna sa stefanocystám pripisoval iný význam, tvrdilo sa, že slúžia k ďalšiemu rozširovaniu huby alebo k pretrvávaniu nepriaznivých podmienok – dôvodom bolo zistenie, že stefanocysty bývajú často odtrhnuté z mycélia a odnášané na tele koristi, prípadne že sa tvorí priamo na bazídiospore.
Sekrečné bunky
Druhov z rodu Pleurotus (napr. Pleurotus ostreatus – hliva ustricová, Pleurotaceae, Basidiomycotina) predstavujú opäť nečakanú a veľmi nezvyčajnú pascu. Sú to malé, oválne, lyžičkovité alebo jazykovité bunky, produkujúce nematotoxín, ktorý na nich vytvára drobné kvapky. Po kontakte koristi s touto látkou dochádza okamžite k jej ochrnutiu – korisť sa okamžite presáva pohybovať, ale zostáva stále živá. Z okolitých hýf predátora začnú rýchlo vyrastať jemné vlákna, smerujúce k telesným otvorom koristi, ktorými ich infikujú. Potom už nasleduje prerastanie asimilačných hýf ďalej do tela koristi a napokon jej strávenie. Niektorý autori považujú tento úplne odlišný typ predácie skôr za parazitizmus. Ekológia týchto drevných húb je veľmi zaujímavá a odvíja sa od limitujúceho faktora v prostredí v ktorom žije – od dusíka.
Toxické hýfy
– huba nemá vytvorenú vôbec žiadnu štruktúru o ktorej by sa dalo tvrdiť, že je to pasca, tou je totižto huba sama. S týmto fenoménom sa môžeme stretnúť iba u niekoľkých druhoch rodu Hypoderma. Potravou tejto huby sa stávajú jedine mykofágne háďatká, ktoré vyhľadávajú hýfy húb, napichujú svojimi stiletami ich bunkové steny a vysávajú obsah. To čo nasleduje je prekvapujúce – pokiaľ si mykofágne háďatko nájde ako potravu hýfy týchto niekoľko druhov húb – ich bunkový obsah je totiž toxický a po jeho pohltení sa predátor stáva korisťou a je znehybnený. Takto ochromenú korisť obopnú hýfy huby a na viacerých miestach ju penetrujú.
Proces chytania koristi
Medzi hubovými predátormi je najrozšírenejší mechanizmus chytania koristi s použitím naviazania lektínov huby na povrchové štruktúry koristi, ale nieje jediný. Rozšírené je tiež napr. i chytanie koristi pomocou buniek produkujúcich toxín, ktorý korisť ochromí a znemožní tým ich únik.
Väčšina lovených organizmov má dobre vyvinuté chemoreceptory (typickým príkladom sú háďatká), ktorými sa riadi pri vyhľadávaní potravy. Tento fakt vedia huby veľmi dobre využiť tým, že produkujú pascami, a v menšej miere i mycéliom, chemické atraktanty, ktoré korisť priťahujú. Akonáhle dôjde ku kontaktu koristi a pasce predátora, nastáva ďalšia fáza.
Na povrchu pasce je prítomné veľké množstvo lektínov, bielkovinových komplexov, ktoré špecificky viažu určité cukry (alebo ich časť, viazané na iné chemické látky). Napr. pasce huby Arthrobotrys oligospora (Deuteromycotina) sú pokryté lektínom, ktorý špecificky viaže N-acetyl D-galaktozamín (GalNAc), ktorý sa v hojnom množstve vyskytuje na kutikule niektorých háďatiek. Hneď po naviazaní väzieb lektín – povrchový uhľovodík je korisť definitívne chytená a sú spustené ďalšie procesy, vedúce k znehybneniu koristi, penetrácii kutikuly a tráveniu. Dôkazy o pôsobení tohto mechanizmu už boli zistené vo väčšom množstve – napr. bol na médium s kolóniou predátora aplikovaný roztok s 20 mM koncentráciou GalNAc. V dôsledku toho došlo k obsadeniu väzobných miest lektínu a vyradenie pascí z prevádzky o čom sa mohli presvedčiť aj háďatká umiestnené do kolónie.
U rôznych húb (a koristi) sa však tieto chemické látky líšia (a zrejme aj rôzne kombinujú) – takže môže dochádzať aj k určitej substrátovej špecifite, ktorá je výrazne vyššia u obligátnych predátorov ako u fakultatívnych. Ako iný príklad tohto mechanizmu môžeme menovať parazitickú hubu Monacrosporium coniospora (Deuteromycotina), čo znamená že je parazit, pretože jeho konídie sú pokryté mukóznou vrstvou obsahujúcou lektíny, ktoré sa preferenčne viažu na senzorické orgány iba niekoľkých rodov háďatiek. V tomto prípade sa špecifické lektíny parazita viažu na kyselinu sialovú, ktorú môžeme nájsť práve iba okolo ústneho otvoru koristi. Je zaujímavé, že nematódy, živiacimi sa baktériami alebo fungivory, sú chytané práve týmto spôsobom, zatiaľ čo rastlinný paraziti sú infikovaní po celom tele. Je možné, že to súvisí s charakterom potravy (nepomer medzi veľkosťou mikrokolónie baktérií alebo hýfou a koreňa) a spôsobom vyhľadávania potravy háďatkom (ktorý je v prvom prípade asi viac cielenejší). Často sa stretávame s prípadmi, kedy podobné alebo rovnaké povrchové látky nachádzame u načisto odlišných organizmov. Príkladom môže byť situácia u Monacrosporium haptotylum (Deuteromycotina), ktorého povrchové lektíny sa špecificky viažu na manán, ktorý je prítomný na kutikule niektorých háďatiek a tiež kvasinky Saccharomyces cerevisiae. V tomto prípade môžu byť alternatívne využívané obidva zdroje, čo je samozrejme výrazná výhoda.
Systém rozoznávania povrchových štruktúr lektínov je medzi týmito hubami a tiež obyčajne v prírode veľmi rozšírený. Nie vždy sa však pri rozoznávaní povrchových štruktúr zúčastňujú iba lektíny. Napr. Monacrosporium ellipsosporum (Deuteromycotina) má pasce pokryté mucín-špecifickým hematoglutenínom s afinitou k niektorým živočíšnym mukopolysacharidom.
Po zachytení koristi nastáva ďalší proces – penetrácie hýfy predátora do tela koristi, čo môže trvať od desiatok minút až do niekoľko hodín. Behom tohto procesu dochádza k narušeniu kutikuly hostiteľa enzýmami, ktoré sa nachádzajú v drobných elektrondenzných telieskach, prítomných v hojnom množstve v cytoplazme pascí. Na ďalšej fáze, kedy dochádza k znehybneniu koristi sa môžu podieľať toxíny, ktoré rovnako obsahuje cytoplazma pasce v drobných inklúziách. Po rozrušení kutikuly a preniknutí hýfy do tela koristi sa často vytvára infekčný bulbus, apresórium, z ktorej vybiehajú tráviace hýfy, pomocou ktorých je korisť v niekoľkých nasledujúcich dňoch celkom rozložená a strávená.
V nasledujúcich videách ponúkame pohľad na predáciu húb pri chytaní koristi a rôzne spôsoby jej chytania.
(Un ventre vide, 2011)
(Un ventre vide, 2011)
(Nandkumar Kamat, 2011)
Taxonómia
Dravé huby sú formálnou skupinou bez akýchkoľvek taxonomických súvislostí. Väčšina doteraz známych zástupcov, ktorých spôsob výživy je možné nazvať pravou predáciou, patrí do oddelenia Eumycota.
Jednu výraznú skupinu nájdeme v triede Zygomycetes, špeciálne v rade Zoopagales. Patria sem zväčša obligátni predátory (poznáme niekoľko desiatok druhov), ktoré môžeme nájsť buď vo vlhkej pôde, komposte, exkrementoch živočíchov a vo vode. Bežne spomínané sú napr. rody Zoopage, Acaulopage, Cystopage, Stylopage a v súčasnosti sa sem zaraďuje i rod Zoophagus, ktorý bol v minulosti zaraďovaný do oddelenia Oomycota a rody Lecophagus, Cephalophora.
Skupinou, v ktorej môžeme nájsť najväčšie množstvo predátorov (ich počet sa pohybuje okolo 100 druhov) je pododdelenie Deuteromycotina. Všetky huby, ktoré môžeme zaradiť do tejto skupiny, sú nepohlavné štádiá (anamorfy), rozmnožujúce sa konídiami. V poslednej dobe boli zistené vzťahy niektorých týchto anamorf k ich teleomorfám (pohlavné štádium, napr. rod Orbilia), ktoré sú určujúce pre zatriedenie do systému a ktoré prislúcha triede Ascomycetes. Vzhľadom k výraznej morfologickej podobnosti zástupcov rodov Arthrobotrys (v súčasnosti sú do nej často zaraďované i dravé huby rodu Dactylaria), Dactylaria a Monacrosporium (ktoré tvoria najväčšiu, najznámejšiu a najviac preskúmanú skupinu dravých húb) a vzhľadom k tomu, že teleomorfné štádium Orbilia bolo nájdené u rôznych druhov, ale aj napriek tomuto môžeme uvažovať, že sú opäť monofyletického pôvodu. Situáciu komplikuje prítomnosť viacerých druhov v tejto skupine, ktoré uplatňujú parazitizmus či saprofytizmus a je možné, že sa u niektorých z nich jedná o druhotnú stratu predácie.
Ako nie príliš súvisiacu zaujímavosť môžeme uviesť búrlivý vývoj systému vo vnútri tejto skupiny húb. V systematike húb sa kľúčovým spôsobom uplatňujú rozdiely v rozmnožovacích štruktúrach. Rovnako tak tomu bolo i v tomto komplexe – rôzne rody boli charakterizované podľa rozdielu v stavbe konídioforov a konídií. Z hľadiska typov pascí však došlo k tomu, že jednotlivé typy pascí sa vyskytovali u rôznych rodov načisto chaoticky a z hľadiska klasifikácie na rodovej úrovni to bolo prakticky nepoužiteľné. Moderné taxonomické štúdie, založené na štúdiu sekvencií ribozomálnej DNA však klasický systém od základu napadli. Ukázalo sa, že skupina by sa mohla rozdeľovať do štyroch skupín, ktoré dobre korešpondujú práve s typom pascí.
V skupine imperfekných húb (Deuteromycotina) môžeme nájsť i ďalšie dravé huby, napr. z rodu Tridentaria, Pedilospora, Triposprina a Haptocara, ktoré sú preskúmané omnoho menej, u niektorých z nich môžeme nájsť isté indície o príbuznosti s predošlou skupinou. Medzi týmito hubami môžeme nájsť ako obligátne, tak i fakultatívne predátory. Väčšinu z nich môžeme nájsť v pôde, hnijúcom dreve a organických zbytkoch, iba výnimočne sú to vodné huby.
Do triedy Homobasidiomycetes, kde je predácia známa asi u 50 druhov, patria 3 nasledujúce rody: Homohenbuehelia (Tricholomataceae) s dravou anamorfou Nematoctonus, Pleurotus (Pleurotaceae) a Hyphoderma (Corticiaceae). V tejto skupine predácia nepochybne vznikla viackrát nezávisle na sebe a existuje podozrenie, že sa vyskytuje i u ďalších rodov (Resupinauts, Stigmatolemma). Všetkých týchto zástupcov nájdeme na mŕtvom, niekedy na silne hnijúcom dreve a patria medzi fakultatívne predátory.
Uvedený stručný prehľad húb predátorov zďaleka nie je kompletný a to hlavne preto, že pri mnohých druhoch je stále obtiažne presne definovať povahu predácie a taktiež očakávať, že u mnohých ďalších tento špecifický spôsob obživy ešte len bude objavený. Mimo toho existujú druhy a rody, ktorých spôsob výživy môžeme bezstarostne označiť ako predáciu, ktoré ale nepatria do pravých húb (Eumycota). Jedná sa napr. o rod Sommerstorffia (druh S. spinosa, Saprolegniales, Oomycota), ktorý sa vyskytuje vo vode. Tento druh na výbežky hýf chytá vírniky a ako alternatívny zdroj energie využíva odumreté stielky rias. Ďalším prípadom je rod Polyphagus (druh P. euglenae, Chytridiales, Chytridiomycota). Táto vodná huba vytvára rhizomycélium (nepravé jedno jadrové mycélium), na ktoré chytá pohybujúce sa alebo encystované bunky rias rodov Euglena a Chlamydomonas a môže v populáciách týchto rias vyvolať epidémiu.
Na záver môžeme povedať, že schopnosť predácie je u húb výnimočná a v priebehu vývoja sa objavila mnohokrát u rôznych skupín húb. Veľmi často nachádzame medzi príbuznými druhmi ako predátory, tak aj druhy uplatňujúce iný spôsob výživy (a len málo predátorov je striktne obligátnych), čo všetko je dôsledkom evolučných a ekologických bariér, brániacich vzniku a širšiemu rozvinutiu predácie v tejto skupine organizmov.
Využitie dravých húb
Záujem o využitie dravých húb sa odvíja od dávno známej skúsenosti, že zvýšenie počtu háďatiek v prostredí (napr. po zelenom hnojení) je nasledované zvýšenou aktivitou hubových predátorov. Bohužiaľ sa ale ukazuje, že na praktické použitie týchto húb v biokontrole parazitických nematód je treba splniť alebo aspoň nejakým spôsobom obísť minimálne tri obtiažne podmienky. Prvou závažnou prekážkou je zaistenie prežitia inokula huby v natívnej populácií húb. Veľmi často dochádza k tzv. fungistatickému efektu, kedy je sťažené klíčenie spór dravých húb a dochádza taktiež k lýze ich klíčnych hýf. Druhou podmienkou je, že huba musí byť schopná zareagovať presne v okamžiku, kedy dochádza k premnoženiu škodcov a tretia je zaistenie prítomnosti a aktivity dravej huby po celú dobu rastu plodín. Je evidentné, že splnenie týchto podmienok nie je jednoduché a hlavne nie je lacné. Aj keď pokusov je veľa, pozitívnych výsledkov je zatiaľ poskromne, mnohokrát potreba použitia ochranného prostriedku s prídavkami dravých húb.
Ako klasické prípady použitia môžeme menovať napr. komerčné prípravky s Arthrobotrys superba a ďalšími druhmi húb proti fytofágnym háďatkám parazitujúcich na koreňoch a iných orgánoch poľnohospodárskych plodín. Napr. Arthrobotrys irregularis je súčasťou prípravku Royal 350, ktorý bol s úspechom použitý proti háďatkám z rodu Meloidogyne na paradajkách a uhorkách. Ako doplnkové látky môžu byť používané niektoré nematocídy a hnojivá (pre zvýšenie vitality rastlín). Ďalej sú dravé huby s úspechom používané (napr. Arthrobotrys superba – ako hlavná zložka prípravku Royal 300) pri pestovaní húb, hlavne šampiňónov, kde môže dôjsť k veľkým škodám spôsobenými mykofágnymi háďatkami, napr. druhom Ditylenchus myceliophagus, ktorý napichuje hýfy pestovaných húb a vysáva ich obsah. V posledných rokoch je s úspechom používaná Duddingtonia (Arthrobotrys) flagrans proti parazitickým črevným hlístam hovädzieho dobytka, oviec a koní. Používajú sa chlamydospóry huby, ktoré sa nechajú prejsť zažívacím traktom zvierat. Potom sa na truse zvierat vytvorí mycélium s pascami, ktoré chytajú larvy parazitov a zabraňujú tak ďalšiemu šíreniu nákazy.
Použitá literatúra:
Barron, G. L. (1992). Lignolytic and cellulolytic fungi as predators and parasites. The fungal community: its organization and role in the ecosystem, 9, 311-326.
Dackman, C., & Nordbring-Hertz, B. (1992). Conidial traps—a new survival structure of the nematode-trapping fungus Arthrobotrys oligospora. Mycological research, 96(3), 194-198.
Dix, N. J. (Ed.). (2012). Fungal ecology. Springer Science & Business Media.
Hawksworth, D. L., Kirk, P. M., Sutton, B. C., & Pegler, D. N. (1995). Dictionary of the fungi (No. C/589.203 H3).
Jaffee, B. A., Strong, D. R., & Muldoon, A. E. (1996). Nematode-trapping fungi of a natural shrubland: tests for food chain involvement. Mycologia, 554-564.
Jensen, C. H., Neumeister, H. G., & Lysek, G. (1997). Nematophagous fungi–study by fluorescence microscopy and EDX-technique of the periodicity of trap formation in soil. Biological Rhythm Research, 28(3), 365-373.
Liou, G. Y., & Tzean, S. S. (1997). Phylogeny of the genus Arthrobotrys and allied nematode-trapping fungi based on rDNA sequences. Mycologia, 876-884.
Pfister, D. H., & Liftik, M. E. (1995). Two Arthrobotrys anamorphs from Orbilia auricolor. Mycologia, 684-688.
Tzean, S. S., & Liou, J. Y. (1993). Nematophagous resupinate basidiomycetous fungi. PHYTOPATHOLOGY-NEW YORK AND BALTIMORE THEN ST PAUL-, 83, 1015-1020.
Webster, J., Henrici, A., & Spooner, B. (1998). Orbilia fimicoloides sp. nov., the teleomorph of Dactylella cf. oxyspora. Mycological Research, 102(1), 99-102.
Citované zdroje z internetu:
Philip Jacobs. 2001. Nematode Pilze. link: http://www.nematophage-pilze.de
Christian Jensen, Heike Neumeister, Gemot Lysek. 1998. Fluorescense microscopy for the observation fungi inside soil. Mycologist, vol. 12, part. 3, pp. 107-111. link: http://www.cjensen.de/html/fluomicro.html
Citované videá z youtube:
Un ventre vide. 2011. Nematophagous fungi part 1. link: https://www.youtube.com/watch?v=14zmmbXsyuM
Un ventre vide. 2011. Nematophagous fungi part 2. link: https://www.youtube.com/watch?v=Uktd10jLPAM
Nandkumar Kamat. 2011. World’s unique battle -Powerful Arthobortrys fungal adhesive employed to capture soil nematode. link: https://www.youtube.com/watch?v=8NPa68QCI-0
Leave a Reply