")
Foto: Michal Kukla antwiki.org
Nikola Hricáková, Lukáš Hleba
Slovenská poľnohospodárska univerzita v Nitre, Fakulta biotechnológia a potravinárstva
Evolúcia húb
Z hľadiska evolúcie predstavujú huby (ríša Fungi) v ekosystéme polyfyletickú skupinu organizmov zúčastňujúcich sa na procese rozkladu organického materiálu a vytváraní pozitívnych a negatívnych interakcií s inými organizmami. Sú jednou z hlavných skupín eukaryotických mikroorganizmov vyskytujúcich sa v pôdnom a vodnom ekosystéme. Kirk a kolektív v roku 2008 uvádzajú, že v súčasnosti existuje približne 100 000 popísaných druhov mikroskopických húb, ktoré zastupujú len časť tejto rozmanitej skupiny, pozostávajúcej odhadom z 1,5 až 5 miliónov druhov.
Vzťahy medzi hubami
Vzťahy medzi hubami a hmyzom môžu byť saprofytické, symbiotické, mutualistické a parazitické. Zvláštnu skupinu však tvoria entomopatogénne huby, ktoré vyvolávajú primárne ochorenie v rôznych vývojových štádiách hmyzu. Vďaka účinným adaptačným mechanizmom sú schopné spustiť proces infekcie v rôznych štádiách vývoja hmyzu a vo väčšine prípadov spôsobujú smrť hostiteľa. Z tohto hľadiska sú zaradené medzi druhy využívané na biologickú ochranu proti škodcom rastlín. Aplikáciu entomopatogénnych húb v biologickej kontrole ovplyvňuje ich schopnosť prispôsobiť sa rozmanitým ekologickým podmienkam prostredia. Zmena abiotických faktorov, predovšetkým teploty, pH, vzdušnej vlhkosti a intenzity slnečného žiarenia, vplýva na schopnosť konídií prežiť a teda aj na schopnosť preniknutia a infikácie hostiteľského organizmu. Veľké množstvo výskumov je sústredených na sledovanie adaptácie entomopatogénnych húb v rozličných biotopoch a na ovplyvnenie kvality infekčného procesu s cieľom zefektívniť aplikáciu entomopatogénov v lesníckej a poľnohospodárskej sfére.
Súčasné taxonomické začlenenie
V súčasnosti je známych viac ako 700 druhov húb zo 100 rodov, ktoré môžu pôsobiť ako obligátni alebo fakultatívni pôvodcovia ochorení mnohých druhov hmyzu. V taxonomickom zaradení sú entomopatogénne druhy zastúpené v kmeňoch Basidiomycota, Chytridiomycota, Oomycota a Zygomycota. Medzi najčastejších hostiteľov patria motýle (Lepidoptera), polokrídlovce (Homoptera), blanokrídlovce (Hymenoptera), chrobáky (Coleoptera) a dvojkrídlovce (Diptera). Najčastejším infikovaným štádiom hmyzu sú larvy, prípadne kukly.
Infekčný cyklus entomopatogénnych húb
Infekčný cyklus entomopatogénnych húb začína penetráciou cez exoskelet a kutikulu hostiteľa. Má dve fázy – parazitickú, počas ktorej dochádza k infekcii hostiteľa a smrti, a saprofytickú, ktorá nastáva po smrti hostiteľa. Vznik a priebeh infekčného procesu ovplyvňuje viacero aspektov. Hlavným sú vlastnosti patogénov a hostiteľa. Prichytenie konídií na povrchu tela hostiteľa je začiatkom ochorenia. Väčšinou dochádza k prenikaniu hubových vláken cez kutikulu hmyzu, menej často sa jedná o penetráciu cez ústne orgány, kontaminovanou potravou, kedy sa spóry dostávajú do tráviaceho traktu a začnú klíčiť. Ďalšou možnosťou je preniknutie infekcie cez pohlavný aparát. Stupeň klíčenia ovplyvňuje ako samotná vnímavosť hostiteľa, tak aj optimálna teplota a relatívna vzdušná vlhkosť. V štádiu klíčenia konídie využívajú zásobné látky, a preto nepotrebujú čerpať živiny z okolia. Pri vysokej relatívnej vzdušnej vlhkosti dochádza z dôvodu prijatia vody k napučaniu konídií a k tvorbe penetračnej štruktúry na špičke klíčka (apresorium), ktorý produkuje enzýmy, ktoré narúšajú štruktúru kutikuly hmyzu. Z apresoria vzniká penetračný hrot, pomocou ktorého infekcia preniká do tela hostiteľa. Po jeho preniknutí nastáva tvorba hýf, ktoré kolonizujú telovú dutinu a dochádza k množeniu pučaním. Patogén produkuje toxíny a sekundárne metabolity, ktorými prekonáva imunitné látky hostiteľa. Parazitická fáza je ukončená smrťou hostiteľa, kedy dochádza ku deštrukcii lymfocytov. Hubové mycélium prerastá na povrch jedinca, dochádza k tvorbe fruktifikačných orgánov. Nasleduje saprofytická fáza, ktorá je ukončená sporuláciou. Prejavom sú nové sekundárne konídie, vitálne niekoľko mesiacov. Životaschopnosť zabezpečuje dormancia konídií, ktorá sa po prichytení na nového jedinca končí.
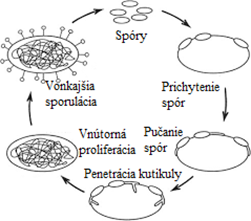
Obrázok č. 1: Typický životný cyklus väčšiny entomopatogénnych húb (upravené podľa Srivastava et al., 2009)
Entomopatogénne druhy
Najväčšie množstvo entomopatogénnych druhov húb sa radí do radu Hypocreales, ktoré sú typické širokým spektrom hostiteľov a tvorbou veľkého množstva sekundárnych metabolitov. Beauveria bassiana, Metarhizium anisopliae radiace sa do radu Hypocreales patria medzi prirodzených nepriateľov škodcov v agroekosystémoch a tým pádom sú vhodnými kandidátmi na budúcu biologickú ochranu do oblasti s miernym podnebím. Prvé pokusy ničenia hmyzích škodcov sa uskutočnili v Rusku, v roku 1888 entomopatogénnou hubou, známou ako Metarhizium anisopliae.
Metarhizium anisopliae
Metarhizium anisopliae (Metchnikoff) Sorokinje anamorfná huba patriaca do čeľade Cordycipitaceae, produkujúca skupinu cyklických peptidových toxínov – destruxínov (DTX), ktoré sú spájané predovšetkým s reguláciou škodcov ako Manduca sexta (Lišaj tabakový), Schistocerca gregaria (Saranča všežravá) a Otiorhynchus sulcatus (Nosánik ryhovaný). Spomínané DTX sú dostatočne silné na to aby zabili hostiteľa skôr, než dôjde k jeho vnútornej proliferácií. Telo hostiteľa je prerastené bielymi hýfami. Po určitej dobe dôjde k zmene farby mycélia na tmavo zelenú v dôsledku vznikajúcich spór. V laboratórnych aj skleníkových podmienkach bola Metarhizium anisopliae testovaná proti Hoplia philantus za spolupôsobenia hlíst Steinernema glaseri a Heterorhabditis megidis. Aplikované boli spoločne s hubovou suspenziou po dobu 2, 3 a 4 týždňov. Na konci výskumu bolo pozorované aditívne a synergistické pôsobenie. Pri ďalšej štúdii bolo pozorované spolupôsobenie hlíst Steinernema carpocapsae a Heterorhabditis bacteriophora a huby Beavueria brongiartii proti Exomala orientalis (Coleoptera). Na zrovnanie bol použitý fenitrothion. Bioagensy mali v porovnaní s chemickou ochranou väčšiu mortalitu.
Isaria fumosorosea
Huba Isaria fumosorosea je testovaná proti zástupcom rodu Diptera, ako Hematobia irritans, Delia radicus a Delia floralis. Účinok bol pozorovaný aj pri zástupcoch Coptotermes formosanus, Trialeurodes vaporariorum.
V roku 1965 v bývalom ZSSR bol vyvinutý mykoinsekticíd, známy ako Boverin. Tento mykoinsekticíd bol vytvorený z entomopatogénnej huby Beauveria bassiana na reguláciu Leptinotarsa decemlineata (Pásavka zemiaková).
Beaveria bassiana
Beaveria bassiana je kozmopolitná vreckatá huba, schopná využívať široký rozsah prostredia, vrátane pôdy, rastlín a hmyzu. Je schopná žiť saprofyticky v pôde, ako endofyt v rastlinách alebo ako entomopatogén v článkonožcoch. Dokáže sa reprodukovať sexuálne v telomorfnom stave nazývanom Cordyceps bassiana. Hmyzí hostitelia pre B. bassiana zahrňujú škodcov plodín (napr. vošky, vijačka kukuričná), ekologicky nebezpečných škodcov (napr. termity) a prenášačov chorôb (komáre, kliešte). Na povrchu infikovaného hostiteľa vytvára husté biele mycélium a preto sa nákazy vyvolané týmito hubami nazývajú ,,biele muskardiny“. Produkuje sekundárny metabolit beauvericín, ktorý oslabuje imunitný systém hostiteľa. Po usmrtení hostiteľa produkuje antibiotikum oosperin, ktoré konkuruje intestinálnym baktériám vo vnútri mŕtveho tela.
Najsilnejšie účinky
Spomedzi zástupcov entomopatogénnych húb práve Beauveria vykazuje najväčšiu virulenciu v boji proti lykožrútovi smrekovému (Ips typographus) avšak aj najväčšiu adaptáciu na tohto hostiteľa v zmysle schopnosti patogéna šíriť sa prostredníctvom mechanizmov odrážajúcich populačné správanie hostiteľa. Najčastejšou formou pôsobenia je aplikácia pomocou spór v podobe vodnej suspenzie na povrch infikovaných stromov. Ďalšou formou je aplikácia práškového koncentrátu spór s cieľom kontaminovať povrch tela hostiteľa a zabezpečiť šírenie nákazy. Využívanie biopreparátov s obsahom Beauveria bassiana je využívané v Rakúsku, Nemecku a experimentálne v Austrálii a USA. Aplikácia sa využíva s cieľom dlhodobejšieho účinku na škodcov.

Obrázok č. 2 Napadnutie Ips typographus (lykožrút smrekový) entomopatogénnou hubou Beauveria bassiana (Vakula et al., 2009)
Zaujímavosť
Zaujímavosťou je, že niektoré druhy hmyzu majú schopnosť detegovať a varovať populáciu na prítomnosť infikovaného jedinca. Veľmi zaujímavé správanie v tomto má sociálny hmyz, ako sú napríklad termity. Prítomnosť infikovaného jedinca v kolónií spôsobí poplach. Po ustúpení poplachu v populácií nasleduje ošetrovanie, kusanie, defekácia a zakopávanie infikovaného jedinca.
Použitá literatúra
ANSARI M.A. – TIRRY L. – MOENS M. 2004. Interaction between Metarhizium anisopliae and entomopathogenic nematodes for the control of Hoplia philanthus. In Biological Control, 2004, č. 31, s. 172 – 180.
AUGUSTYNIUK-KRAM, A. – KRAM, K.J. 2012. Entomopathogenic fungi as an important natural regulator of insect outbreaks in forests. In Blanco, J.A., Yueh-Hsin, L. Forest ecosystems – More than just trees. Croatia : Intech, p. 265-295. ISBN 978-953-51-0202-1.
CHARNLEY, A. K. (1997). Entomopathogenic fungi and their role in pest control. The Mycota IV: Environmental and microbial relationships, 185-201.
CHOO H.Y. – KAYA H.K. – HUH J. – LEE D.W. – KIMM H.H. – LEE S.M. – CHOO Y.M., 2002. Entomopathogenic nematodes (Steinenema spp. and Heterorhabditis bacteriophora) and fungus Beauveria brongniarti for biological control of white grubs, Ectinopholia rufipes and Exomala orientalis, In Korean golf course. BioControl, 2002, č. 47, s. 177 – 192.
INGLIS G.D. et al. 2001: Use of Hyphomycetes fungi for manging Insect Pests. In Butt T.M., Jackson C., Magan N. Fungi as biocontrol agents – progress, problems and potential. Wallingford, UK : CAB International, pp. 23-69. ISBN 0-85199-356-7.
JARONSKI, S.T. 2010. Ecological factors in the inundative use of fungal entomopathogens. In Biological Control 2010, vol. 55, pp. 159–185. ISSN 1049-9644KERSHAW, M. J. et al.. (1999). The role of destruxins in the pathogenicity of Metarhizium anisopliae for three species of insect. Journal of invertebrate pathology, 74(3), 213-223.
KIRK, P.M. et al. 2008. Dictionary of the fungi. 10th Edition. Wallingford, UK : CAB International. 640p. ISBN 9780643095731.
KOUBOVÁ, D. (2009). Využití hub v biologické ochraně rostlin proti škůdcům.
LANDA, Z. – KŘENOVÁ, Z. – VOJTĚCH, O. 2007. Využití houby Beauveria bassiana v ochraně proti lýkožroutu smrkovému. Lesnická práce, s. 14-15.
MEYLING, N.V. – EILENBERG, J. (2007). Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: potential for conservation biological control. Biological control, 43(2), 145-155.
MUELLER, G.M. – SCHMIT, J.P. 2007. Fungal biodiversity: what do we know? What can we predict? In Biodiversity and Conservation, vol. 16, pp. 1-5. ISSN 1572-9710.
OSBORNE, L.S. – LANDA, Z. 1992. Biological Control of Whiteflies with Entomopathogenic Fungi. In Florida Entomologist, vol. 75, no. 4, pp. 456-471. ISSN 1938-5102.
SAMSON , R.A. – EVANS, H.C. – LATGE, J.P. 1988. Atlas of entomopathogenic fungi. Germany : Springer. 187p. ISBN 978-3-662-05890-9.
SCHOLTE, K. – KNOLS, R. – TAKKEN, W. (2008). An entomopathogenic fungus (Metarhizium anisopliae) for control of the adult African malaria vector Anopheles gambiae. ENTOMOLOGISCHE BERICHTEN-NEDERLANDSCHE ENTOMOLOGISCHE VEREENIGUNG, 68(1), 21.
SRIVASTAVA, CH.N. et al. 2009. Prospective role of insecticides of fungal origin: Review. In Entomological research, vol. 39, no. 6, pp. 341-355. ISSN 1748-5967.
VALERO-JIMENÉZ, C. A., et al. (2016). Genes involved in virulence of the entomopathogenic fungus Beauveria bassiana. Journal of invertebrate pathology, 133, 41-49.
VÄNNINEN I. – HOKKANEN H. – TYNI-JUSLIN J. 1999. Attempts to control cabbage root flies Delia radicum L. and Delia floral (Fall.) (Dipt., Anthomyiidae) with entomopathogenic fungi:laboratory and greenhouse tests. In Journal of Applied Entomology, 1999, č. 123, s. 107 – 113.
VEY, A. – HOAGLAND, B. – BUTT, T. M. 2001. Toxic metabolites of fungal biocontrol agents. In Butt, T., Jackson, C. W., Magan, N. Fungi as biocontrol agents: progress, problems and potential, Wallington : CAB International, 2001, pp. 311-346. ISBN 0-85199-356-7.
ZIMMERMANN,G. 2007. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. In Biocontrol Science and Technology, vol. 17, pp. 553–596. ISSN 2456-5717. zȩcS
Leave a Reply